
Y – DNA – Haplogroup R1b – Our McKee Haplogoup
Author – Maciamo Hay in Eupedia.com/genetics – adapted by Barry McKee
Contents
- Paleolithic Mammoth Hunters
- Neolithic Cattle Herders
- The Levantine and African Branch (V88)
- The Pontic-Caspian Steppe – the Indo-European Link
- The Maykop Culture, the R1b Link to the Steppe?
- R1b Migration Map
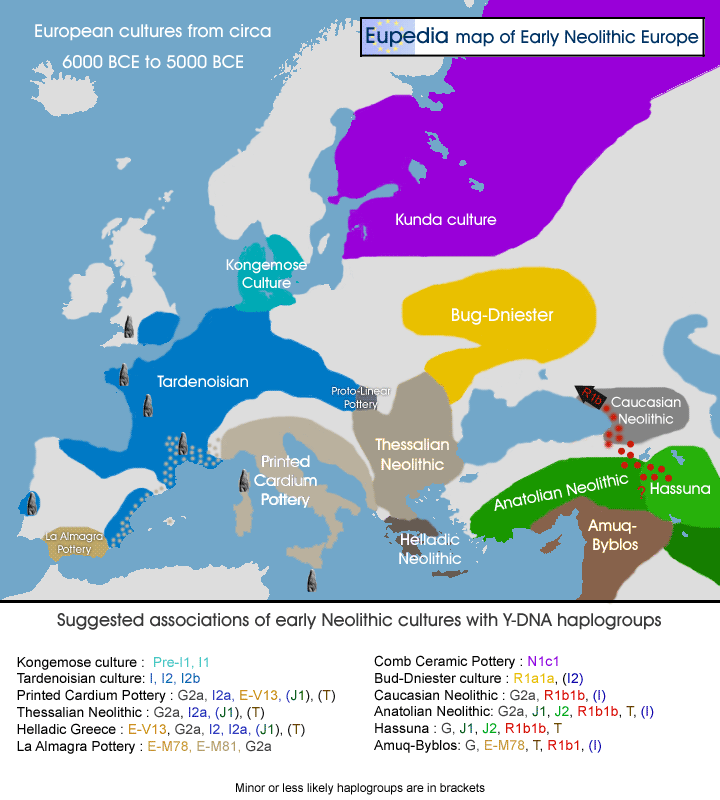
- Early Neolithic Cultures with Associated Haplogroups
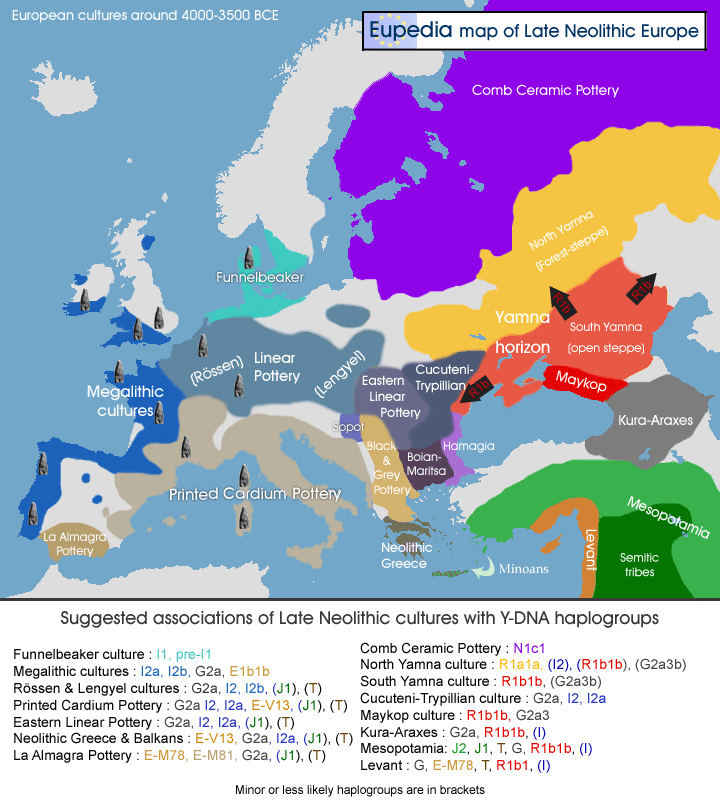
- Late Neolithic Cultures with Associated Haplogroups
- The Siberian and Central Asian Branch
- The European and Middle Eastern Branch
- The Conquest of “Old Europe” and Central Europe
- The Conquest of Western Europe
- Metal-Mining and Stockbreeding Explain R1b Dominance in Atlantic Fringe
- Did the Indo-Europeans Really Invade Western Europe?
- The Atlantic Celtic Branch (L21)
- The Gascon-Iberian Branch (DF27)
- The Italo-Celtic Branch (S28/U152)
- The Germanic Branch (S21/U106)
- How Did R1b Become Dominant ?
- The Balkan and Asian Branch (Z2103)
- The Upheavals circa 1200 BCE
- Other Migrations of R1b
- Lactase Persistence and R1b Cattle Pastoralists
- R1 Populations & Light Pigmentation
- The Maternal Lineages (mtDNA) Corresponding to Haplogroup R1b
Disclaimer This information about R1b Haplogroup should not be read as hard facts, but, as is often the case in science, as a model in constant evolution based on the present knowledge and understanding (of the author, Maciamo Hay). Whenever the advancement of genetics couldn’t provide irrefutable answers, he has attempted to provide the most likely and logical hypothesis based on archeological, historical and linguistic evidence.
1. Geographic Distribution of Haplogroup R1b
Distribution of Haplogroup R1b in Europe
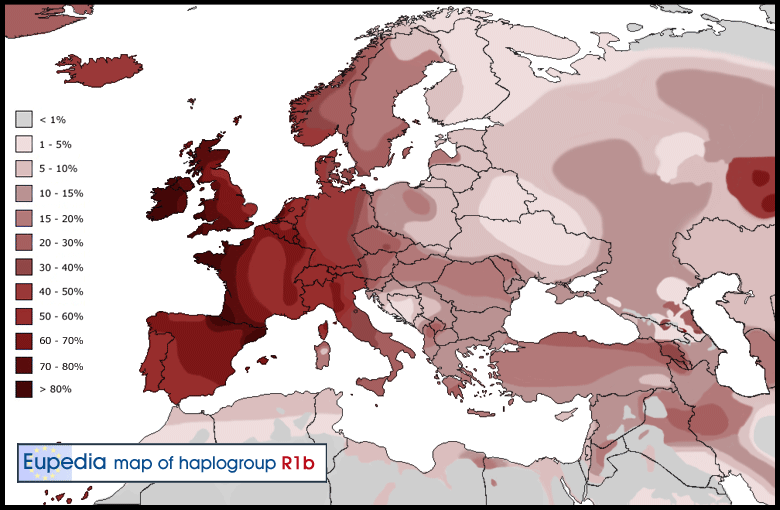
R1b (our McKee Family Haplogroup) is the most common haplogroup in Western Europe, reaching over 80% of the population in Ireland, the Scottish Highlands, western Wales, the Atlantic fringe of France, the Basque country and Catalonia. It is also common in Anatolia and around the Caucasus, in parts of Russia and in Central and South Asia. Besides the Atlantic and North Sea coast of Europe, hotspots include the Po valley in north-central Italy (over 70%), Armenia (35%), the Bashkirs of the Urals region of Russia (50%), Turkmenistan (over 35%), the Hazara people of Afghanistan (35%), the Uyghurs of North-West China (20%) and the Newars of Nepal (11%). R1b-V88, a subclade specific to sub-Saharan Africa, is found in 60 to 95% of men in northern Cameroon.
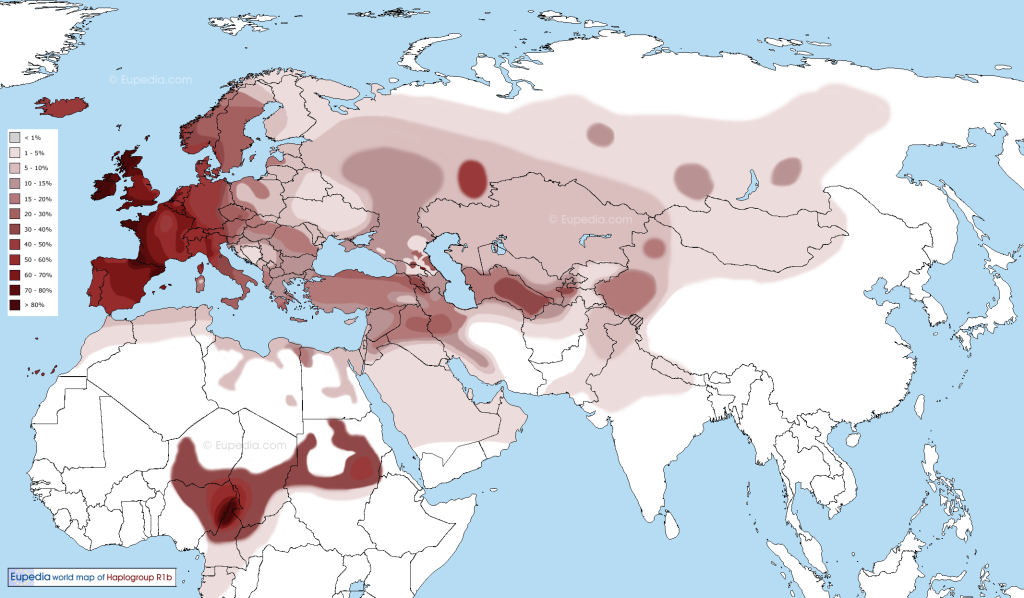
Distribution Map of Haplogroup R1b in the Old World
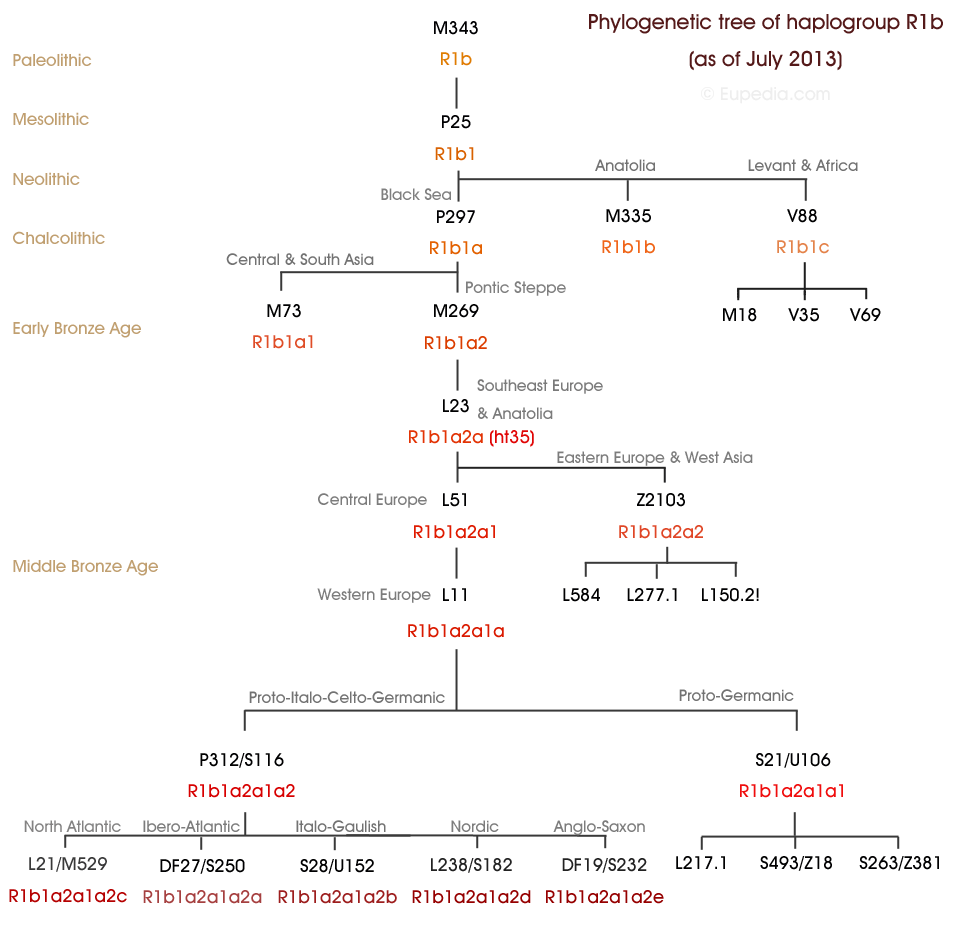
2. Subclades (Subgroups) of Haplogroup R1b
Here is a schematic tree of the principal R1b Subclades (subgroups). My (our) McKee subgroup is L21 at the lower left of the tree. Refer to the International Society of Genetic Genealogy (ISOGG) for the full tree with all the SNP’s and the latest nomenclature.If you are not familiar with the time-scale at the left of this chart, see the timeline in Wikipedia http://en.wikipedia.org/wiki/Timeline_of_human_prehistory for the prehistory period (Paleolithic, Mesolithic (with their boundary at 25,000 years before now) and Neolithic (10,000 to 5,500 years ago). Chalcolithic refers to the Copper age, about 7000 – 6000 years ago (or 5000 – 4000 years BCE).
3. Origins and History of Haplogroup R1b
Paleolithic Mammoth Hunters
Haplogroup R* originated in North Asia just before the Last Glacial Maximum (26,500-19,000 years ago). This haplogroup has been identified in the remains of a 24,000 year-old boy from the Altai region, in south-central Siberia (Raghavan et al. 2013). This individual belonged to a tribe of mammoth hunters that may have roamed across Siberia and parts of Europe during the Paleolithic. With Autosomal DNA results, this Paleolithic population appears to have contributed mostly to the ancestry of modern Europeans and South Asians, the two regions where haplogroup R also is the most common nowadays (R1b in Western Europe, R1a in Eastern Europe, Central and South Asia, and R2 in South Asia).
The oldest forms of R1b (M343, P25, L389) are found dispersed at very low frequencies from Western Europe to India, a vast region where the nomadic R1b hunter-gatherers could have roamed during the Ice Age. The three main branches of R1b1 (R1b1a, R1b1b, and R1b1c) all seem to have stemmed from the Middle East. The southern branch, R1b1c (V88), is found mostly in the Levant and Africa. The northern branch, R1b1a (P297), seems to have originated around the Caucasus, eastern Anatolia or northern Mesopotamia, and then to have crossed over the Caucasus, from where they would have invaded Europe and Central Asia. R1b1b (M335) has only been found in Anatolia.
Neolithic Cattle Herders
It has been hypothesized that R1b people (perhaps alongside neighbouring J2 tribes) were the first to domesticate cattle in northern Mesopotamia some 10,500 years ago. R1b tribes descended from mammoth hunters, and when mammoths went extinct, they started hunting other large game such as bisons and aurochs. With the increase of the human population in the Fertile Crescent from the beginning of the Neolithic (starting 12,000 years ago), selective hunting and culling of herds started replacing indiscriminate killing of wild animals. The increased involvement of humans in the life of aurochs, wild boars and goats led to their progressive taming. Cattle herders probably maintained a nomadic or semi-nomadic existence, while other people in the Fertile Crescent (presumably represented by haplogroups E1b1b, G and T) settled down to cultivate the land or keep smaller domesticates.
The analysis of bovine DNA has revealed that all the taurine cattle (Bos taurus) alive today descend from a population of only 80 aurochs. The earliest evidence of cattle domestication dates from circa 8,500 BCE in the Pre-Pottery Neolithic cultures in the Taurus Mountains. The two oldest archaeological sites showing signs of cattle domestication are the villages of Çayönü Tepesi in southeastern Turkey and Dja’de el-Mughara in northern Iraq, two sites only 250 km away from each other. This is presumably the area from which R1b lineages started expanding – or in other words the “original homeland” of R1b. The early R1b cattle herders would have split in at least three groups. One branch (M335) remained in Anatolia, but judging from its extreme rarity today wasn’t very successful, perhaps due to the heavy competition with other Neolithic populations in Anatolia, or to the scarcity of pastures in this mountainous environment. A second branch migrated south to the Levant, where it became the V88 branch. Some of them searched for new lands south in Africa, first in Egypt, then colonising most of northern Africa, from the Mediterranean coast to the Sahel. The third branch (P297), crossed the Caucasus into the vast Pontic-Caspian Steppe, which provided ideal grazing grounds for cattle. They split into two factions: R1b1a1 (M73), which went east along the Caspian Sea to Central Asia, and R1b1a2 (M269), which at first remained in the North Caucasus and the Pontic Steppe between the Dnieper and the Volga. It is not yet clear whether M73 actually migrated across the Caucasus and reached Central Asia via Kazakhstan, or if it went south through Iran and Turkmenistan. In the latter case, M73 might not be an Indo-European branch of R1b, like V88 and M335.
R1b-M269 (the most common form in Europe) is closely associated with the diffusion of Indo-European languages, as attested by its presence in all regions of the world where Indo-European languages were spoken in ancient times, from the Atlantic coast of Europe to the Indian subcontinent, which comprised almost all Europe (except Finland, Sardinia and Bosnia-Herzegovina), Anatolia, Armenia, European Russia, southern Siberia, many pockets around Central Asia (notably in Xinjiang, Turkmenistan, Tajikistan and Afghanistan), without forgetting Iran, Pakistan, northern India and Nepal. The history of R1b and R1a are intricately connected to each others.
The Levantine and African Branch of R1b (V88)
Like its northern counterpart (R1b-M269), R1b-V88 is associated with the domestication of cattle in northern Mesopotamia. Both branches of R1b probably split soon after cattle were domesticated, approximately 10,500 years ago (8,500 BCE). R1b-V88 migrated south towards the Levant and Egypt. The migration of R1b people can be followed archeologically through the presence of domesticated cattle, which appear in central Syria around 8,000-7,500 BCE (late Mureybet period), then in the Southern Levant and Egypt around 7,000-6,500 BCE (e.g. at Nabta Playa and Bir Kiseiba). Cattle herders subsequently spread across most of northern and eastern Africa. The Sahara desert would have been more humid during the Neolithic Subpluvial period (c. 7250-3250 BCE), and would have been a vast savannah full of grass, an ideal environment for cattle herding.
Evidence of cow herding during the Neolithic has shown up at Uan Muhuggiag in central Libya around 5500 BCE, and at the Capeletti Cave in northern Algeria around 4500 BCE. But the most compelling evidence that R1b people related to modern Europeans once roamed the Sahara is to be found at Tassili n’Ajjer in southern Algeria, a site famous for pyroglyphs (rock art) dating from the Neolithic era. Some painting dating from around 3000 BCE depict fair-skinned and blond or auburn haired women riding on cows.
After reaching the Maghreb, R1b-V88 cattle herders could have crossed the Strait of Gibraltar to Iberia, probably accompanied by G2 farmers, J1 and T1a goat herders and native Maghreban E-M81 lineages. These Maghreban Neolithic farmers/herders could have been the ones who established the Almagra Pottery culture in Andalusia in the 6th millennium BCE. Nowadays small percentages (1 to 4%) of R1b-V88 are found in the Levant, among the Lebanese, the Druze, and the Jews, and almost in every country in Africa north of the equator. Higher frequency in Egypt (5%), among Berbers from the Egypt-Libya border (23%), among the Sudanese Copts (15%), the Hausa people of Sudan (40%), the Fulani people of the Sahel (54% in Niger and Cameroon), and Chadic tribes of northern Nigeria and northern Cameroon (especially among the Kirdi), where it is observed at a frequency ranging from 30% to 95% of men. According to Cruciani et al. (2010) R1b-V88 would have crossed the Sahara between 9,200 and 5,600 years ago, and is most probably associated with the diffusion of Chadic languages, a branch of the Afro-asiatic languages. V88 would have migrated from Egypt to Sudan, then expanded along the Sahel until northern Cameroon and Nigeria. However, R1b-V88 is not only present among Chadic speakers, but also among Senegambian speakers (Fula-Hausa) and Semitic speakers (Berbers, Arabs).
R1b-V88 is found among the native populations of Rwanda, South Africa, Namibia, Angola, Congo, Gabon, Equatorial Guinea, Ivory Coast, and Guinea-Bissau. The wide distribution of V88 in all parts of Africa, its incidence among herding tribes, and the coalescence age of the haplogroup all support a Neolithic dispersal. In any case, a later migration out of Egypt would be improbable since it would have brought haplogroups that came to Egypt during the Bronze Age, such as J1, J2, R1a or R1b-L23. The maternal lineages associated with the spread of R1b-V88 in Africa are mtDNA haplogroups J1b, U5 and V, and perhaps also U3 and some H subclades.
The North Caucasus and the Pontic-Caspian Steppe : the Indo-European Language Link
Modern linguists have placed the Proto-Indo-European homeland in the Pontic-Caspian Steppe, a distinct geographic and archeological region extending from the Danube estuary to the Ural mountains to the east and North Caucasus to the south. The Neolithic, Chalcolithic and early Bronze Age cultures in the Pontic-Caspian steppe has been called the Kurgan culture (4200-2200 BCE) by Marija Gimbutas, due to the lasting practice of burying the dead under mounds (“kurgan”) among the succession of cultures in that region. It is now known that kurgan-type burials only date from the 4th millennium BCE and almost certainly originated south of the Caucasus. The genetic diversity of R1b being greater around eastern Anatolia, it is hard to deny that R1b evolved there before entering the steppe world. Horses were first domesticated around 4600 BCE in the Caspian Steppe, perhaps somewhere around the Don or the lower Volga, and soon became a defining element of steppe culture. Nevertheless it is unlikely that R1b was already present in the eastern steppes at the time, so the domestication of the horse should be attributed to the indigenous R1a people.
It is not yet entirely clear when R1b crossed over from eastern Anatolia to the Pontic-Caspian steppe. This might have happened with the appearance of the Dnieper-Donets culture (c. 5100-4300 BCE). This was the first truly Neolithic society in the Pontic-Caspian steppe. Domesticated animals (cattle, sheep and goats) were herded throughout the steppes and funeral rituals were elaborate. Sheep wool would play an important role in Indo-European society, notably in the Celtic and Germanic (R1b branches of the Indo-Europeans) clothing traditions up to this day. However, many elements indicate a continuity in the Dnieper-Donets culture with the previous Mesolithic hunter-gatherers, and at the same time an influence from the Balkans and Carpathians, with regular imports of pottery and copper objects. It is therefore more likely that Dnieper-Donets marked the transition of indigenous R1a and/or I2a1b people to early agriculture, perhaps with an influx of Near Eastern farmers from ‘Old Europe’. Mitochondrial DNA sequences from Dnieper-Donets culture showed clear similarities with those of the Cucuteni-Trypillian culture in the Carpathians (haplogroups H, T and U3).
The first clearly Proto-Indo-European culture was Sredny Stog (4600-3900 BCE), when small kurgan burials begin to appear, with the distinctive posturing of the dead on the back with knees raised and oriented toward the northeast, which would be found in later steppe cultures as well. There is evidence of population blending from the variety of skull shapes. Towards the end of the 5th millennium, an elite starts to develop with cattle, horses and copper used as status symbols.
Another migration across the Caucasus happened shortly before 3700 BCE, when the Maykop culture, the world’s first Bronze Age society, suddenly materialized in the north-west Caucasus, apparently out of nowhere. The origins of Maykop are still uncertain, but archeologists have linked it to contemporary Chalcolithic cultures in Assyria and western Iran. Archeology also shows a clear diffusion of bronze working and kurgan-type burials from the Maykop culture to the Pontic Steppe, where the Yamna culture developed soon afterwards (from 3500 BCE). Kurgan (a.k.a. tumulus) burials would become a dominant feature of ancient Indo-European societies and were widely used by the Celts, Romans, Germanic tribes, and Scythians, among others.
The Yamna period (3500-2500 BCE) is the most important one in the creation of Indo-European culture and society. Middle Eastern R1b people had been living and blending to some extent with the local R1a foragers and herders for over a millennium, perhaps even two or three. The close cultural contact and interactions between R1a and R1b people all over the Pontic-Caspian Steppe resulted in the creation of a common vernacular, a new lingua franca, which linguists have called Proto-Indo-European (PIE). It is pointless to try to assign another region of origin to the PIE language. Linguistic similarities exist between PIE and Caucasian and Hurrian languages in the Middle East on the one hand, and Uralic languages in the Volga-Ural region on the other hand, which makes the Pontic Steppe the perfect intermediary region.
During the Yamna period cattle and sheep herders adopted wagons to transport their food and tents, which allowed them to move deeper into the steppe, giving rise to a new mobile lifestyle that would eventually lead to the great Indo-European migrations. This type of mass migration in which whole tribes moved with the help of wagons was still common in Gaul at the time of Julius Caesar, and among Germanic peoples in the late Antiquity.
The Yamna horizon was not a single, unified culture. In the south, along the northern shores of the Black Sea coast until the north-west Caucasus, was a region of open steppe, expanding eastward until the Caspian Sea, Siberia and Mongolia (the Eurasian Steppe). The western section, between the Don and Dniester Rivers (and later the Danube), was the one most densely settled by R1b people, with only a minority of R1a people (5-10%). The eastern section, in the Volga basin until the Ural mountains, was inhabited by R1a people with a substantial minority of R1b people (whose descendants can be found among the Bashkirs, Turkmans, Uyghurs and Hazaras, among others). The northern part of the Yamna horizon was forest-steppe occupied by R1a people, also joined by a small minority of R1b (judging from modern Russians and Belarussians, the frequency of R1b was from seven to nine times less lower than R1a). The western branch would migrate to the Balkans and Greece, then to central and Western Europe, and back to their ancestral Anatolia in successive waves (Hittites, Phrygians, Armenians, etc.). The eastern branch would migrate to Central Asia, Xinjiang, Siberia, and South Asia (Iran, Pakistan, India). The northern branch would evolve into the Corded Ware culture and disperse around the Baltic, Poland, Germany and Scandinavia.
The Maykop Culture, the R1b Link to the Steppe ?
The Maykop culture (3700-2500 BCE) in the north-west Caucasus was culturally speaking a sort of southern extension of the Yamna horizon. Although not generally considered part of the Pontic-Caspian steppe culture due to its geography, the North Caucasus had close links with the steppes, as attested by numerous ceramics, gold, copper and bronze weapons and jewelry in the contemporaneous cultures of Mikhaylovka, Sredny Stog and Kemi Oba. The link between the northern Black Sea coast and the North Caucasus is older than the Maykop period. Its predecessor, the Svobodnoe culture (4400-3700 BCE), already had links to the Suvorovo-Novodanilovka and early Sredny Stog cultures. The even older Nalchik settlement (5000-4500 BCE) in the North Caucasus displayed a similar culture as Khvalynsk on the Volga. This may be the period when R1b started interacting and blending with the R1a population of the steppes.
The Yamna and Maykop people both used kurgan burials, placing their dead in a supine position with raised knees and oriented in a north-east/south-west axis. Graves were sprinkled with red ochre on the floor, and sacrificed domestic animal buried alongside humans. They also had in common horses, wagons, a heavily cattle-based economy with a minority of sheep kept for their wool, use of copper/bronze battle-axes (both hammer-axes and sleeved axes) and tanged daggers. In fact, the oldest wagons and bronze artefacts are found in the North Caucasus, and appear to have spread from there to the steppes.
Maykop was an advanced Bronze Age culture, actually one of the very first to develop metalworking, and therefore metal weapons. The world’s oldest sword was found at a late Maykop grave. Its style is reminiscent of the long Celtic swords, though less elaborate. Horse bones and depictions of horses already appear in early Maykop graves, suggesting that the Maykop culture might have been founded by steppe people or by people who had close links with them. However, the presence of cultural elements radically different from the steppe culture in some sites could mean that Maykop had a hybrid population. Without DNA testing it is impossible to say if these two populations were an Anatolian R1b group and a G2a Caucasian group, or whether R1a people had settled there too. The two or three ethnicities might even have cohabited side by side in different settlements. The one typical Caucasian Y-DNA lineage that does follow the pattern of Indo-European migrations is G2a3b1, which is found throughout Europe, Central Asia and South Asia. In the Balkans, the Danube basin and Central Europe its frequency is somewhat proportional to the percentage of R1b.
Maykop people are the ones credited for the introduction of primitive wheeled vehicles (wagons) from Mesopotamia to the steppes. This would revolutionise the way of life in the steppe, and would later lead to the development of (horse-drawn) war chariots around 2000 BCE. Cavalry and chariots played an vital role in the subsequent Indo-European migrations, allowing them to move quickly and defeat easily anybody they encountered. Combined with advanced bronze weapons and their sea-based culture, the western branch (R1b) of the Indo-Europeans from the Black Sea shores are excellent candidates for being the mysterious Sea Peoples, who raided the eastern shores of the Mediterranean during the second millennium BCE.
The rise of the IE-speaking Hittites in Central Anatolia happened a few centuries after the disappearance of the Maykop and Yamna cultures. Considering that most Indo-European forms of R1b found in Anatolia today belong to the R1b-Z2103 subclade, there is little doubt that the Hittites came to Anatolia via the Balkans, after Yamna/Maykop people invaded Southeast Europe. The Maykop and Yamna cultures were succeeded by the Srubna culture (1600-1200 BCE), possibly representing an advance of R1a1a people from the northern steppes towards the Black Sea shores, filling the vacuum left by the R1b tribes who migrated to Southeast Europe and Anatolia.
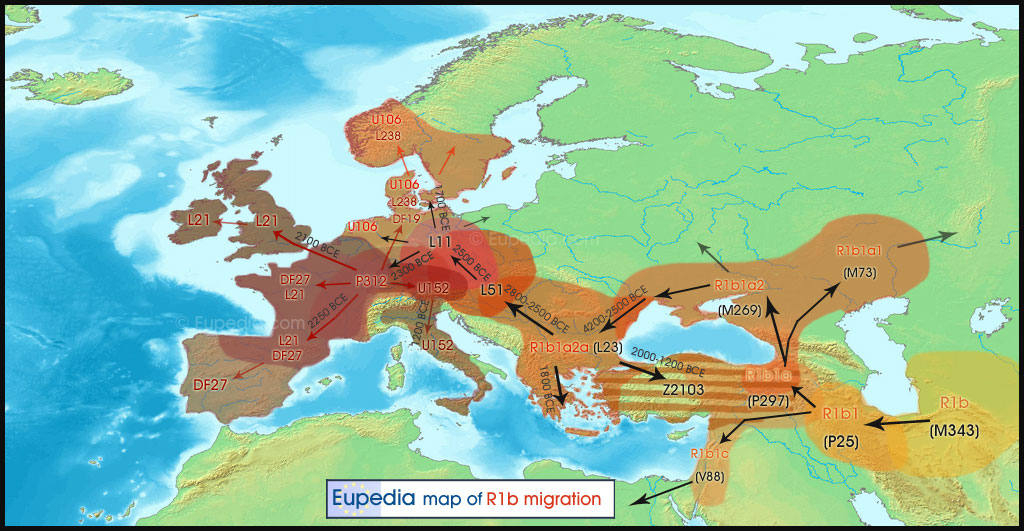
Migration Map of Y-Haplogroup R1b from the Paleolithic to the End of the Bronze Age – c. 1000 BCE
Map of Early Neolithic Cultures Associated with Y-DNA Haplogroups — 6000 – 5000 BCE (below)
Map of Late Neolithic Cultures Associated with Y-DNA Haplogroups — 4000 – 3500 BCE (below)
The Siberian and Central Asian branch
When R1b crossed the Caucasus in the Late Neolithic, it split into two main groups. The western one (L51) would settle the eastern and northern of the Black Sea. The eastern one (Z2103 + M73) migrated to the Don-Volga region, where horses were domesticated circa 4600 BCE. R1b probably mixed with indigenous R1a people and founded the Repin culture (3700-3300 BCE) a bit before the Yamna culture came into existence in the western Pontic Steppe. R1b would then have migrated with horses along the Great Eurasian Steppe until the Altai mountains in East-Central Asia, where they established the Afanasevo culture (c. 3600-2400 BCE). Afanasevo people might be the precursors of the Tocharian branch of Indo-European languages alongside haplogroup R1a.
The R1b people who stayed in the Volga-Ural region were probably the initiators of the Poltavka culture (2700-2100 BCE), then became integrated into the R1a-dominant Sintashta-Petrovka culture (2100-1750 BCE) linked to the Indo-Aryan conquest of Central and South Asia (=> see R1a for more details).
Nowadays R1b is found at higher frequencies among ethnic minorities of the Volga-Ural region in Russia than among Slavic Russians. R1b is also present in many Central Asian populations, the highest percentages being observed among the Uyghurs (20%) of Xinjiang in north-west China, the Yaghnobi people of Tajikistan (32%), and the Bashkirs (47%, or 62.5% in the Abzelilovsky district) of Bashkortostan in Russia (border of Kazakhstan).
It is not entirely clear at present whether R1b1a1 (M73), found primarily in North Asia (Altai, Mongolia), Central Asia and the North Caucasus is of Indo-European or pre-Indo-European origin. It could have spread from West Asia around the Caspian Sea and north to Siberia and Mongolia during the Neolithic. But alternatively it could have been an Indo-European lineage also found in the Volga-Ural region, alongside Z2103. In that scenario, M73 could have spread to the Altai region during the Afanasevo period (perhaps together with R1a lineages).
The European and Middle Eastern branch
The Indo-Europeans’ bronze weapons and the extra mobility provided by horses would have given them a tremendous advantage over the autochthonous inhabitants of Europe, namely the native haplogroup I (descendant of Cro-Magnon), and the early Neolithic herders and farmers (G2a, J, E1b1b and T). This allowed R1a and R1b to replace most of the native male lineages, although female lineages seem to have been less affected.
A comparison with the Indo-Iranian invasion of South Asia shows that 40% of the male linages of northern India are R1a, but less than 10% of the female lineages could be of Indo-European origin. The impact of the Indo-Europeans was more severe in Europe because European society 4,000 years ago was less developed in terms of agriculture, technology (no bronze weapons) and population density than that of the Indus Valley civilization. This is particularly true of the native Western European cultures where farming arrived much later than in the Balkans or Central Europe. Greece, the Balkans and the Carpathians were the most advanced of European societies at the time and were the least affected in terms of haplogroup replacement. Native European Y-DNA haplogroups (I1, I2) also survived better in regions that were more difficult to reach or less hospitable, like Scandinavia, southern Switzerland, Sardinia or the Dinaric Alps.
The Conquest of “Old Europe” and Central Europe (4200-2500 BCE)
The first forays of steppe people into the Balkans happened between 4200 BCE and 3900 BCE, when cattle herders equipped with horse-drawn wagons crossed the Dniester and Danube and apparently destroyed the towns of the Gumelnita, Varna and Karanovo VI cultures in Eastern Romania and Bulgaria. A climatic change resulting in colder winters during this exact period probably pushed steppe herders to seek milder pastures for their stock, while failed crops would have led to famine and internal disturbance within the Danubian and Balkanic communities. The ensuing Cernavoda culture (Copper Age, 4000-3200 BCE), Coțofeni culture (Copper to Bronze Age, 3500-2500 BCE) and Ezero culture (Bronze Age, 3300-2700 BCE), in modern Romania, seems to have had a mixed population of steppe immigrants and people from the old tell settlements. These steppe immigrants were likely a mixture of both R1a and R1b lineages, with a probably higher percentage of R1a than later Yamna-era invasions.
The steppe invaders would have forced many Danubian farmers to migrate to the Cucuteni-Trypillian towns in the eastern Carpathians, causing a population boom and a north-eastward expansion until the Dnieper valley, bringing Y-haplogroups G2a, I2a1 (probably the dominant lineage of the Cucuteni-Trypillian culture), E1b1b, J and T in what is now central Ukraine. This precocious Indo-European advance westward was fairly limited, due to the absence of Bronze weapons and organised army at the time, and was indeed only possible thanks to climatic catastrophes which reduced the defences of the towns of Old Europe. The Carpathian, Danubian, and Balkanic cultures were too densely populated and technologically advanced to allow for a massive migration.
In comparison, the forest-steppe R1a people successfully penetrated into the heart of Europe with little hindrance, due to the absence of developed agrarian societies around Poland and the Baltic. The Corded Ware (Battle Axe) culture (3200-1800 BCE) was a natural western expansion of the Yamna culture, reaching as far west as Germany and as far north as Sweden and Norway. DNA analysis from the Corded Ware culture site of Eulau confirms the presence of R1a (but not R1b) in central Germany around 2600 BCE. The Corded Ware tribes expanded from the northern fringe of the Yamna culture where R1a lineages were prevalent over R1b ones.
The expansion of R1b people into Old Europe was slower, but proved inevitable. In 2800 BCE, by the time the Corded Ware had already reached Scandinavia, the Bronze Age R1b cultures had barely moved into the Pannonian steppe. They established major settlements in the Great Hungarian Plain, the most similar habitat to their ancestral Pontic Steppes. Around 2500 BCE, the western branch of Indo-European R1b were poised for their next major expansion into modern Germany and Western Europe. By that time, the R1b immigrants had blended to a great extent with the indigenous Mesolithic and Neolithic populations of the Danubian basin, where they had now lived for 1,700 years.
The strongly patriarchal Indo-European elite remained almost exclusively R1b on the paternal side, but absorbed a high proportion of non-Indo-European maternal lineages. Hybridised, the new Proto-Indo-European R1b people would have lost most of their remaining Proto-Europoid or Mongolid features inherited from their Caspian origins (which were still clearly visible in numerous individuals from the Yamna period). Their light hair, eye and skin pigmentation, once interbred with the darker inhabitants of Old Europe, became more like that of modern Southern Europeans. The R1a people of the Corded Ware culture would come across far less populous societies in Northern Europe, mostly descended from the lighter Mesolithic population (haplogroup I1 and I2), and therefore retain more of their original pigmentation (although facial traits evolved considerably in Scandinavia, where the I1 inhabitants were strongly dolicocephalic and long-faced, as opposed to the brachycephalic and broad-faced steppe people).
The Conquest of Western Europe (2500-1200 BCE)
The R1b conquest of Europe happened in two phases. For nearly two millennia, starting from circa 4200 BCE, steppe people limited their conquest to the rich Chalcolithic civilisations of the Carpathians and the Balkans. These societies possessed the world’s largest towns, notably the tell settlements of the Cucuteni-Tripolye culture. Nothing incited the R1b conquerors to move further into Western Europe at such an early stage, because most of the land north and west of the Alps was still sparsely populated woodland. The Neolithic did not reach the British Isles and Scandinavia before circa 4000 BCE. Even northern France and most of the Alpine region had been farming or herding for less than a millennium and were still quite primitive compared to Southeast Europe and the Middle East.
North-west Europe remained a tribal society of hunter-gatherers practising only limited agriculture for centuries after the conquest of the Balkans by the Indo-Europeans. Why would our R1b “conquistadors” leave the comfort of the wealthy and populous Danubian civilisations for the harsh living conditions that lie beyond ? Bronze Age people coveted tin, copper, and gold, of which the Balkans had plenty, but that no one had yet discovered in Western Europe.
R1b-L51 is thought to have arrived in Central Europe (Hungary, Austria, Bohemia) around 2500 BCE, approximately two millennia after the shift to the Neolithic in these regions. Agrarian towns had started to develop. Gold and copper had begun to be mined. The prospects of a conquest were now far more appealing.
The archeological and genetic evidence (distribution of R1b subclades) point at several consecutive waves towards eastern and central Germany between 2800 BCE and 2300 BCE. The Unetice culture was probably the first culture in which R1b-L11 lineages played a major role. It is interesting to note that the Unetice period happen to correspond to the end of the Maykop (2500 BCE) and Kemi Oba (2200 BCE) cultures on the northern shores of the Black Sea, and their replacement by cultures descended from the northern steppes. It can therefore be envisaged that the (mostly) R1b population from the northern half of the Black Sea migrated westward due to pressure from other Indo-European people (R1a) from the north, for example that of the burgeoning Proto-Indo-Iranian branch, linked to the contemporary Poltavka and Abashevo cultures.
It is doubtful that the Bell Beaker culture (2800-1900 BCE) in Western Europe was already Indo-European because its attributes are in perfect continuity with the native Megalithic cultures. The Beaker phenomenon started during the Late Neolithic and Early Chalcolithic in Portugal and propagated to the north-east towards Germany. During the same period Bronze Age steppe cultures spread from Germany in the opposite direction towards Iberia, France and Britain. It is more likely that the beakers and horses found across Western Europe during that period were the result of trade with neighbouring Indo-European cultures, including the first wave of R1b into Central Europe. It is equally possible that the Beaker people were R1b merchants or explorers who travelled across Western Europe and brought back tales of riches poorly defended by Stone Age people and waiting to be conquered by the more advanced Indo-Europeans, with their bronze weapons and horses.
What is undeniable is that the following Unetice (2300-1600 BCE), Tumulus (1600-1200 BCE), Urnfield (1300-1200 BCE) and Hallstatt (1200-750 BCE) cultures were linked to the diffusion of R1b to Europe, as they abruptly introduce new technologies and a radically different lifestyle.
Metal-Mining and Stockbreeding Explain R1b Dominance in Atlantic Fringe
About one year ago, I realized that haplogroup R1b originated somewhere between Central Asia and the Middle East, then moved through the Pontic steppe where it became associated with Indo-European culture, before pushing its way through the Danube valley and Western Europe. It all made sense. The older subclades of R1b were found East. R1b1a was found among the Levantines and R1b1* in central Africa with a trail leading back to Egypt and the Middle East 12,000-15,000 years ago. The highest diversity of R1b1b1 and R1b1b2 was found in Anatolia and the Caucasus, and the split between these two haplogroups occurred around Anatolia. Both subclades were found in Russia, Central Asia and Pakistan, an obvious sign that they had been part of the Indo-European migrations, at least in Asia. One thing kept bugging me with this nice theory. How did R1b lineages come to replace most of the older lineages in Western Europe ? I tried to explain that with a series of factors (polygamy, status, war…), but somehow that still didn’t explain why R1b reached tremendous frequencies in places like Ireland, Wales, Cornwall, Brittany or the Basque country, and not elsewhere. I think I have come up with a reasonable answer to this mystery. I was looking at a map of metal-rich zones in Europe, and more specifically where copper, tin, silver and gold mines had been established in the Copper and Bronze Age. The richest regions were the Anatolia, North Caucasus, the Carpathians (Romania), the Balkans (especially central Bulgaria), the Alps, and the Atlantic fringe of Europe ( Ireland, Wales, Cornwall, Brittany). This was exactly the migration route I had established a year ago for R1b1b2. R1b people were evidently metal workers.
The second element that dawned on me is that the Atlantic fringe from Galicia to the Scottish Highlands, must have been poor agricultural land for early farmers. This may be why farming spread later to these regions, and that aboriginal Megalithic cultures thrived there more than anywhere else. What does that have to do with R1b ? The Proto-Italo-Celts that brought R1b lineages to Western Europe from the North Caucasus and Pontic steppes had an economy relying on stockbreeding and herding more than farming. The European Bronze Age is characterized by the sharp diminution in agriculture and an increase in domesticates. The steppe culture was replacing the Neolithic lifestyle. Where else could this have the most dramatic effect on the population structure than in the Atlantic fringe, where the Neolithic/Chalcolithic population was sparser than elsewhere due to the late adoption of agriculture and low yield of the farms?
The arrival of the metal-hungry, horse-riding and warlike Indo-Europeans with their bronze swords and axes was a death sentence to the locals. The green pastures of the Atlantic were a boon for the flocks of cattle and herds of sheep of the Indo-Europeans. It was like a milder-climate version of the steppes with the added bonus of being rich in copper and tin, the two components of bronze.
Did the Indo-Europeans Really Invade Western Europe?
Proponents of the Paleolithic or Neolithic continuity model argue that bronze technology and horses could have been imported by Western Europeans from their Eastern European neighbours, and that no actual Indo-European invasion need be involved. It is harder to see how Italic, Celtic and Germanic languages were adopted by Western and Northern Europeans without at least a small scale invasion. It has been suggested that Indo-European (IE) languages simply disseminated through contact, just like technologies, or because it was the language of a small elite and therefore its adoption conferred a certain perceived prestige. However people don’t just change language like that because it sounds nicer or more prestigious. Even nowadays, with textbooks, dictionaries, compulsory language courses at school, private language schools for adults and multilingual TV programs, the majority of the people cannot become fluent in a completely foreign language, belonging to a different language family. The linguistic gap between pre-IE vernaculars and IE languages was about as big as between modern English and Chinese. English, Greek, Russian and Hindi are all related IE languages and therefore easier to learn for IE speakers than non-IE languages like Chinese, Arabic or Hungarian. From a linguistic point of view, only a wide-scale migration of IE speakers could explain the thorough adoption of IE languages in Western Europe – leaving only Basque as a remnant of the Neolithic languages.
One important archeological argument in favour of the replacement of Neolithic cultures by Indo-European culture in the Bronze Age comes from pottery styles. The sudden appearance of bronze technology in Western Europe coincides with ceramics suddenly becoming more simple and less decorated, just like in the Pontic steppes. Until then, pottery had constantly evolved towards greater complexity and details for over 3,000 years. People do not just decide like that to revert to a more primitive style. Perhaps one isolated tribe might experiment with something simpler at one point, but what are the chances that distant cultures from Iberia, Gaul, Italy and Britain all decide to undertake such an improbable shift around the same time ? The best explanation is that this new style was imposed by foreign invaders. In this case it is not mere speculation; there is ample evidence that this simpler pottery is characteristic of the steppes associated with the emergence of Proto-Indo-European speakers.
Besides pottery, archeology provides ample evidence that the early Bronze Age in Central and Western Europe coincides with a radical shift in food production. Agriculture experiences an abrupt reduction in exchange for an increased emphasis on domesticates. This is also a period when horses become more common and cow milk is being consumed regularly. The overall change mimics the steppe way of life almost perfectly. Even after the introduction of agriculture around 5200 BCE, the Bug-Dniester culture and later steppe cultures were characterized by an economy dominated by herding, with only limited farming. This pattern expands into Europe exactly at the same time as bronze working.
Religious beliefs and arts undergo a complete reversal in Bronze Age Europe. Neolithic societies in the Near East and Europe had always worshipped female figurines as a form of fertility cult. The steppe cultures, on the contrary, did not manufacture female figurines. As bronze technology spreads from the Danube valley to Western Europe, symbols of fertility and fecundity progressively disappear and are replaced by cultures of domesticated animals.
Another clue that Indo-European steppe people came in great number to Central and Western Europe is to be found in burial practices. Neolithic Europeans either cremated their dead (e.g. Cucuteni-Tripolye culture) or buried them in collective graves (this was the case of Megalithic cultures). In the steppe, each person was buried individually, and high-ranking graves were placed in a funeral chamber and topped by a circular mound. The body was typically accompanied by weapons (maces, axes, daggers), horse bones, and a dismantled wagon (or later chariot). These characteristic burial mounds are known as kurgans in the Pontic steppe. Men were given more sumptuous tombs than women, even among children, and differences in hierarchy are obvious between burials. The Indo-Europeans had a strongly hierarchical and patrilineal society, as opposed to the more egalitarian and matrilineal cultures of Old Europe. The proliferation of status-conscious male-dominant kurgans (or tumulus) in Central Europe during the Bronze Age is a clear sign that the ruling elite had now become Indo-European. The practice also spread to central Asia and southern Siberia, two regions where R1a and R1b lineages are found nowadays, just like in Central Europe. The ceremony of burial is one of the most emotionally charged and personal aspect of a culture. It is highly doubtful that people would change their ancestral practice “just to do like the neighbours”. In fact, different funerary practices have co-existed side by side during the European Neolithic and Chalcolithic. The ascendancy of yet another constituent of the Pontic steppe culture in the rest of Europe, and in this case one that does not change easily through contact with neighbours, adds up to the likelihood of a strong Indo-European migration. The adoption of some elements of a foreign culture tends to happen when one civilization overawes the adjacent cultures by its superiority. This process is called ‘acculturation’. However there is nothing that indicates that the steppe culture was so culturally superior as to motivate a whole continent, even Atlantic cultures over 2000 km away from the Pontic steppes, to abandon so many fundamental symbols of their own ancestral culture, and even their own language. In fact, Old Europe was far more refined in its pottery and jewellery than the rough steppe people. The Indo-European superiority wasn’t cultural but military, thanks to horses, bronze weapons and an ethic code valuing individual heroic feats in war (these ethic values are known from the old IE texts, like the Rig Veda, Avesta, or the Mycenaean and Hittite literature)
After linguistics and archeology, the third category of evidence comes from genetics itself. It had first been hypothesized that R1b was native to Western Europe, because this is where it was most prevalent. It has since been proven that R1b haplotypes displayed higher microsatellite diversity in Anatolia and in the Caucasus than in Europe. European subclades are also more recent than Middle Eastern or Central Asian ones. The main European subclade, R-P312/S116, only dates back to approximately 3500 to 3000 BCE. It does not mean that the oldest common ancestor of this lineage arrived in Western Europe during this period, but that the first person who carried the mutation R-P312/S116 lived at least 5,000 years ago, assumedly somewhere in the lower Danube valley or around the Black Sea. In any case this timeframe is far too recent for a Paleolithic origin or a Neolithic arrival of R1b. The discovery of what was thought to be “European lineages” in Central Asia, Pakistan and India hit the final nail on the coffin of a Paleolithic origin of R1b in Western Europe, and confirmed the Indo-European link.
All the elements concur in favour of a large scale migration of Indo-European speakers (possibly riding on horses) to Western Europe between 2500 to 2100 BCE, contributing to the replacement of the Neolithic or Chalcolithic lifestyle by a inherently new Bronze Age culture, with simpler pottery, less farming, more herding, new rituals (single graves) and new values (patrilineal society, warrior heroes) that did not evolve from local predecessors.
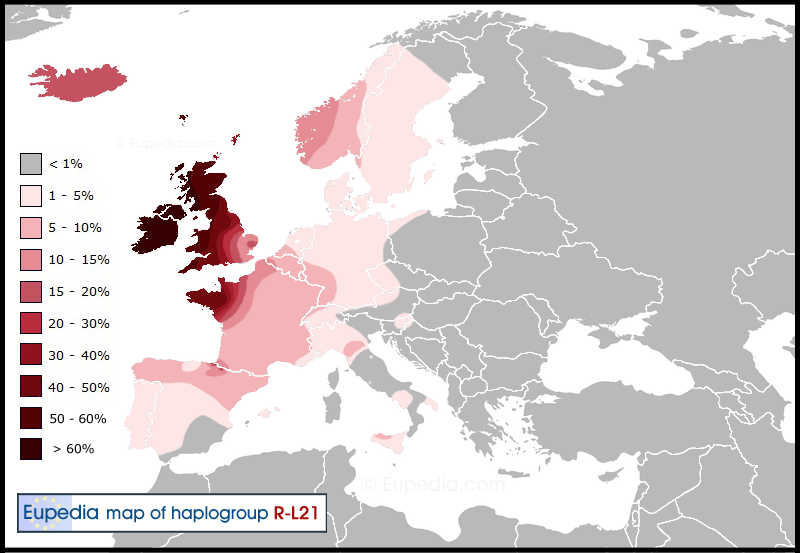
The Atlantic Celtic Branch (L21) Our McKee Branch
The Proto-Italo-Celto-Germanic R1b people had settled in what is now Germany by 2300 BCE, where they founded the Unetice culture. Judging from the propagation of bronze working to Western Europe, those first Indo-Europeans reached France and the Low Countries by 2200 BCE, Britain by 2100 BCE and Ireland by 2000 BCE, and Iberia by 1800 BCE. This first wave of R1b presumably carried R1b-L21 lineages in great number (perhaps because of a founder effect), as these are found everywhere in western, northern and Central Europe. The early split of L21 from the main Proto-Celtic branch around Germany would explain why the Q-Celtic languages (Goidelic and Hispano-Celtic) diverged so much from the P-Celtic branch (La Tène, Gaulish, Brythonic), which appears to have expanded from the later Urnfield and Hallstat cultures.
Some L21 lineages from the Netherlands and northern Germany later entered Scandinavia (from 1700 BCE) with the dominant subclade of the region, R1b-S21/U106 (see below). The stronger presence of L21 in Norway and Iceland can be attributed to the Norwegian Vikings, who had colonised parts of Scotland and Ireland and taken slaves among the native Celtic populations, whom they brought to their new colony of Iceland and back to Norway. Nowadays about 20% of all Icelandic male lineages are R1b-L21 of Scottish or Irish origin.
In France, R1b-L21 is mainly present in historical Brittany (including Maylene and Vendee) and in Lower Normandy. This region was repopulated by massive immigration of insular Britons in the 5th century due to pressure from the invading Anglo-Saxons. However, it is possible that L21 was present in Armorica since the Bronze age or the Iron age given that the tribes of the Armorican Confederation of ancient Gaul already had a distinct identity from the other Gauls and had maintained close ties with the British Isles at least since the Atlantic Bronze Age.
Distribution of haplogroup R1b-L21 (S145) in Europe
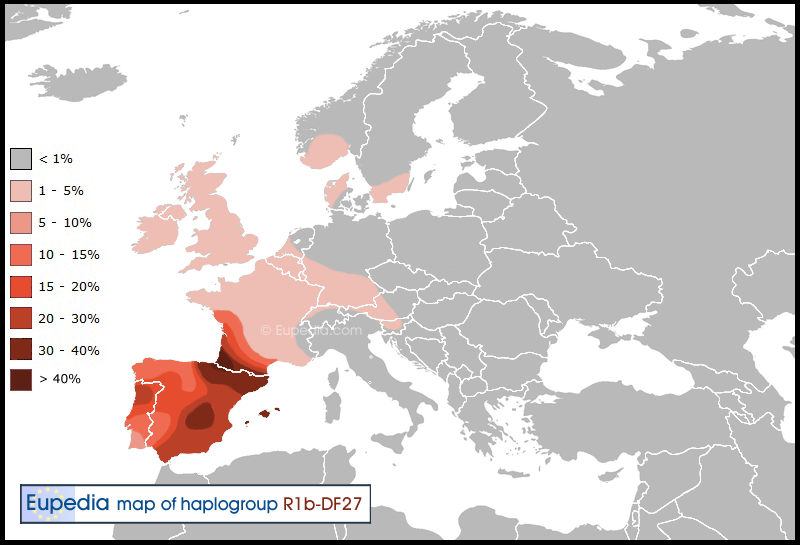
The Gascon and Iberian branch (DF27)
The Bronze Age did not appear in Iberia until 1800 BCE, and was mostly confined to the cultures of El Argar and Los Millares in south-east Spain, with sporadic sites showing up in Castile by 1700 BCE and in Extremadura and southern Portugal by 1500 BCE. These Early Bronze Age sites typically did not have more than some bronze daggers or axes and cannot be considered proper Bronze Age societies, but rather Copper Age societies with occasional bronze artefacts (perhaps imported). These cultures might have been founded by small groups of R1b adventurers looking for easy conquests in parts of Europe that did not yet have bronze weapons. They would have become a small ruling elite, would have had children with local women, and within a few generations their Indo-European language would have been lost, absorbed by the indigenous languages.
Iberia did not become a fully-fledged Bronze Age society until the 13th century BCE, when the Urnfield culture (1300-1200 BCE) expanded from Germany to Catalonia via southern France, then the ensuing Hallstatt culture (1200-750 BCE) spread throughout most of the peninsula (especially the western half). This period belongs to the wider Atlantic Bronze Age (1300-700 BCE), when Iberia was connected to the rest of Western Europe through a complex trade network.
It is hard to say when exactly DF27 entered Iberia. Considering its overwhelming presence in the peninsula and in south-west France, it is likely that DF27 arrived early, during the 1800 to 1300 BCE period, and perhaps even earlier, if R1b adventurers penetrated the Bell Beaker culture, as they appear to have done all over Western Europe from 2300 BCE to 1800 BCE. The Atlantic Bronze Age could correspond to the period when DF27 radiated more evenly around Iberia and ended up, following Atlantic trade routes, all the way to the British Isles, the Netherlands and western Norway (where M153 and SRY2728 make up about 1% of the population).
Distribution of haplogroup R1b-DF27 in Europe
The Italo-Celtic branch (S28/U152)
Starting circa 1300 BCE, a new Bronze Age culture flourished around the Alps thanks to the abundance of metal in the region, and laid the foundation for the classical Celtic culture. It was actually the succession of three closely linked culture: the Urnfield culture, which would evolve into the Hallstatt culture (from 1200 BCE) and eventually into the La Tène culture (from 450 BCE). After the Unetice expansion to Western Europe between 2300 and 1800 BCE, the Urnfield/Hallstatt/La Tène period represents the second major R1b expansion that took place from Central Europe, pushing west to the Atlantic, north to Scandinavia, east to the Danubian valley, and eventually as far away as Greece, Anatolia, Ukraine and Russia, perhaps even to the Tarim basin in north-west China.
The Celtic Iron Age (late Halstatt, from 800 BCE) may have been brought through preserved contacts with the steppes and the North Caucasus, notably the Koban culture (1100-400 BCE).
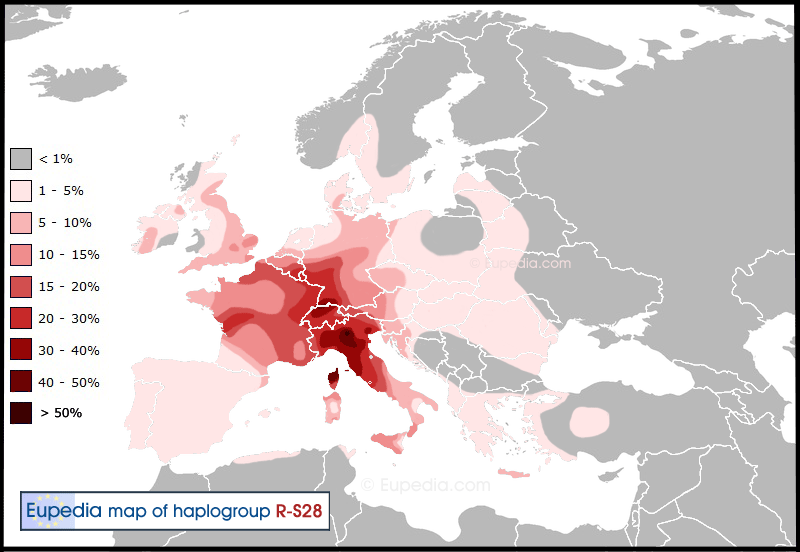
The Alpine Celts of the Hallstatt culture are associated with the S28 (a.k.a. U152) mutation, although not exclusively. The Italic branch (also S28/U152) is thought to have entered Italy in the 12th century BCE (see Villanova culture below), but there were certainly several successive waves, as attested by the later arrival of the Cisalpine Celts from Gaul. The Belgae were another S28/U152 branch, an extension of the La Tène culture northward, following the Rhine, Moselle and Meuse rivers.
One common linguistic trait between Italic and Gaulish/Brythonic Celtic languages linked to the Hallstatt expansion is that they shifted the original IE *kw sound into *p. They are known to linguists as the P-Celtic branch (as opposed to Q-Celtic). It is thought that this change occurred due to the inability to pronounce the *kw sound by the pre-Indo-European population of Central Europe, Gaul and Italy, who were speakers of Afro-Asiatic dialects that had evolved from Near-Eastern languages inherited from the Neolithic. The Etruscans, although later incomers from the eastern Mediterranean, also fit in this category. It has recently been acknowledged that Celtic languages borrowed part of their grammar from Afro-Asiatic languages. This shift could have happened when the Proto-Italo-Celtic speakers moved from the steppes to the Danube basin and mixed with the population of Near-Eastern farmers belonging to haplogroups E1b1b, G2a, J and T. However, such an early shift would not explain why Q-Celtic and Germanic languages did not undergo the same linguistic mutation. It is therefore more plausible that the shift happened after the Proto-Italo-Celts and Proto-Germanics had first expanded across all western and northern Europe. The S28/U152 connection to P-Celtic (and Italic) suggests that the shift took place around the Alps after 1800 BCE, but before the invasion of Italy by the Italic tribes circa 1200 BCE.
The expansion of the Urnfield/Hallstatt culture to Italy is evident in the form of the Villanovan culture (c. 1100-700 BCE), which shared striking resemblances with the Urnfield/Hallstatt sites of Bavaria and Upper Austria. The Villanova culture marks a clean break with the previous Terramare culture. Although both cultures practised cremation, whereas Terramare people placed cremated remains in communal ossuaries like their Neolithic ancestors from the Near East, Villanovans used distinctive Urnfield-style double-cone shaped funerary urns, and elite graves containing jewellery, bronze armour and horse harness fittings were separated from ordinary graves, showing for the first time the development of a highly hierarchical society, so characteristic of Indo-European cultures.
Quintessential Indo-European decorations, such as swastikas, also make their appearance. Originally a Bronze-age culture, the Villanova culture introduced iron working to the Italian peninsula around the same time as it appeared in the Hallstatt culture, further reinforcing the link between the two cultures. In all likelihood, the propagation of the Villanova culture represents the Italic colonisation of the Italian peninsula. The highest proportion of R1b-S28 is found precisely where the Villanovans were the more strongly established, around modern Tuscany and Emilia-Romagna. The Villanova culture was succeeded by the Etruscan civilisation, which displayed both signs of continuity with Villanova and new hybrid elements of West Asian origins, probably brought by Anatolian settlers (who would have belonged to a blend of haplogroups E1b1b, G2a, J1, J2 and R1b-L23).
Distribution of Haplogroup R1b-S28 (U152) in Europe
The Germanic Branch (S21/U106)
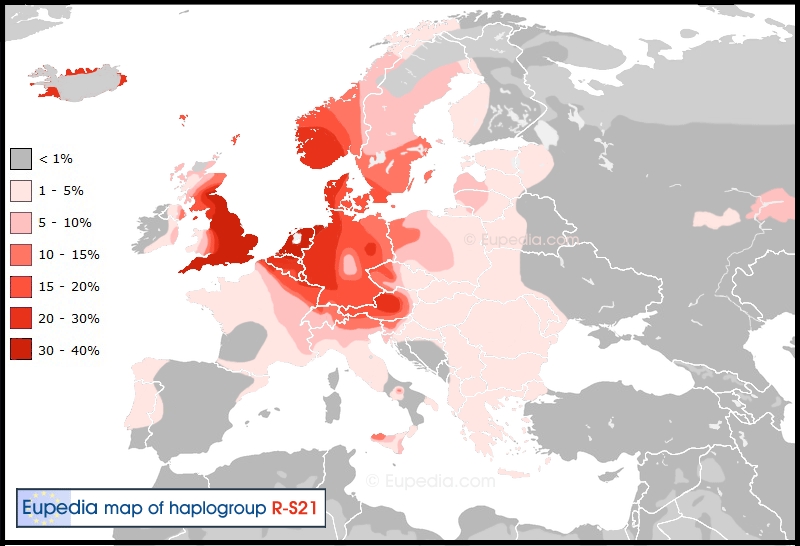
The principal Proto-Germanic branch of the Indo-European family tree is R1b-S21 (a.k.a. U106). This haplogroup is found at high concentrations in the Netherlands and north-west Germany. It is likely that R1b-S21 lineages expanded in this region through a founder effect during the Unetice period, then penetrated into Scandinavia around 1700 BCE, thus creating a new culture, that of the Nordic Bronze Age (1700-500 BCE). R1b-S21 would then have blended for more than a millennium with pre-existing Scandinavian populations, represented by haplogroups I1, I2-M223, R1a-Z284 and to a lesser extent N1c1, which evolved into a relatively unified whole during the Iron Age, the first truly Germanic culture and language, although spread across many tribes. R1b-S21 became the dominant haplogroup among the West Germanic tribes, but remained in the minority against I1 and R1a in East Germanic tribes, including those originating from Sweden such as the Goths, the Vandals and Lombards.
The presence of R1b-S21 in other parts of Europe can be attributed almost exclusively to the Germanic migrations that took place between the 3rd and the 10th century. The Frisians and Anglo-Saxons disseminated this haplogroup to England and the Scottish Lowlands, the Franks to Belgium and France, the Burgundians to eastern France, the Suebi to Galicia and northern Portugal, and the Lombards to Austria and Italy. The Goths help propagate S21 around Eastern Europe, but apparently their Germanic lineages were progressively diluted by blending with Slavic and Balkanic populations, and their impact in Italy, France and Spain was very minor. Later the Danish and Norwegian Vikings have also contributed to the diffusion of R1b-S21 (alongside I1, I2b1 and R1a) around much of Western Europe, but mainly in Iceland, in the British Isles, in Normandy, and in the southern Italy.
From the Late Middle Ages until the early 20th century, the Germans expanded across much of modern Poland, pushing as far as Latvia to the north-east and Romania to the south-east. During the same period the Austrians built an empire comprising what is now the Czech Republic, Slovakia, Hungary, Slovenia, Croatia, Serbia, and parts of Romania, western Ukraine and southern Poland. Many centuries of German and Austrian influence in central and Eastern Europe resulted in a small percentage of Germanic lineages being found among modern populations. In Romania 4% of the population still consider themselves German. The low percentage of R1b-S21 in Finland, Estonia and Latvia can be attributed to the Swedish or Danish rule from the late Middle Ages to the late 19th century.
Distribution of Haplogroup R1b-S21 (U106) in Europe
How did R1b Come to Replace Most of the Older Lineages in Western Europe ?
Until recently it was believed that R1b originated in Western Europe due to its strong presence in the region today. The theory was that R1b represented the Paleolithic Europeans (Cro-Magnon) that had sought refuge in the Franco-Cantabrian region at the peak of the last Ice Age, then recolonized Central and Northern Europe once the ice sheet receded. The phylogeny of R1b proved that this scenario was not possible, because older R1b clades were consistently found in Central Asia and the Middle East, and the youngest in Western and Northern Europe. There was a clear gradient from East to West tracing the migration of R1b people (see map above). This age of the main migration from the shores of the Black Sea to Central Europe also happened to match the timeframe of the Indo-European invasion of Europe, which coincides with the introduction of the Bronze-Age culture in Western Europe, and the proliferation of Italo-Celtic and Germanic languages.
Historians and archeologists have long argued whether the Indo-European migration was a massive invasion, or rather a cultural diffusion of language and technology spread only by a small number of incomers. The answer could well be “neither”. Proponents of the diffusion theory would have us think that R1b is native to Western Europe, and R1a alone represent the Indo-Europeans. The problem is that haplogroup R did arise in Central Asia, and R2 is still restricted to Central and South Asia, while R1a and the older subclades of R1b are also found in Central Asia. The age of R1b subclades in Europe coincide with the Bronze-Age. R1b must consequently have replaced most of the native Y-DNA lineages in Europe from the Bronze-Age onwards.
However, a massive migration and nearly complete annihilation of the Paleolithic population can hardly be envisaged. Western Europeans do look quite different in Ireland, Holland, Aquitaine or Portugal, despite being all regions where R1b is dominant. Autosomal DNA studies have confirmed that the Western European population is far from homogeneous. A lot of maternal lineages (mtDNA) also appear to be of Paleolithic origin (e.g. H1, H3, U5 or V) based on ancient DNA tests. What a lot of people forget is that there is also no need of a large-scale exodus for patrilineal lineages to be replaced fairly quickly. Here is why:
Polygamy. Unlike women, men are not limited in the number of children they can procreate. Men with power typically have more children. This was all the truer in primitive societies, where polygamy was often the norm for chieftains and kings.
Status and Power. Equipped with Bronze weapons and horses, the Indo-Europeans would have easily subjugated the Neolithic farmers and with even greater ease Europe’s last hunter-gatherers. If they did not exterminate the indigenous men, the newcomers would have become the new ruling class, with a multitude of local kings, chieftains and noblemen (Bronze-Age Celts and Germans lived in small village communities with a chief, each part of a small tribe headed by a king) with higher reproductive opportunities than average.
Gender imbalance. Invading armies normally have far more men than women. Men must therefore find women in the conquered population. Wars are waged by men, and the losers suffer heavier casualties, leaving more women available to the winners.
Aggressive warfare. The Indo-Europeans were a warlike people with a strong heroic code emphasising courage and military prowess. Their superior technology (metal weapons, wheeled vehicles drawn by horses) and attitude to life would have allowed them to slaughter any population that did not have organised armies with metal weapons (i.e. anybody except the Middle-Eastern civilizations).
Genetic predisposition to conceive boys. The main role of the Y-chromosome in man’s body is to create sperm. Haplogroups are determined based on mutations differentiating Y-chromosomes. Each mutation is liable to affect sperm production and sperm motility. Preliminary research has already established a link between certain haplogroups and increased or reduced sperm motility. The higher the motility, the higher the chances of conceiving a boy. It is absolutely possible that R1b could confer a bias toward more male offspring. Even a slightly higher percentage of male births would significantly contribute to the replacement of other lineages with the accumulation effect building up over a few millennia. Not all R1b subclades might have this boy bias. The bias only exist in relation to other haplogroups found in a same population. It is very possible that the fairly recent R1b subclades of Western Europe had a significant advantage compared to the older haplogroups in that region, notably haplogroup I2 and E-V13.
Replacement of patrilineal lineages following this model quickly becomes exponential. Imagine 100 Indo-European men conquering a tribe of 1000 indigenous Europeans (a ratio of 1:10). War casualties have resulted in a higher proportion of women in the conquered population. Let’s say that the surviving population is composed of 700 women and 300 men. Let’s suppose that the victorious Indo-European men end up having twice as many children reaching adulthood as the men of the vanquished tribe. There is a number of reason for that. The winners would take more wives, or take concubines, or even rape women of the vanquished tribe. Their higher status would guarantee them greater wealth and therefore better nutrition for their offspring, increasing the chances of reaching adulthood and procreating themselves. An offspring ratio of 2 to 1 for men is actually a conservative estimate, as it is totally conceivable that Bronze-Age sensibilities would have resulted in killing most of the men on the losing side, and raping their women (as attested by the Old Testament). Even so, it would only take a few generations for the winning Y-DNA lineages to become the majority. For instance, if the first generation of Indo-Europeans had two surviving sons per man, against only one per indigenous man, the number of Indo-European paternal lineages would pass to 200 individuals at the second generation, 400 at the third, 800 at the fourth and 1600 at the fifth, and so on. During that time indigenous lineages would only stagnate at 300 individuals for each generation.
Based on such a scenario, the R1b lineages would have quickly overwhelmed the local lineages. Even if the Indo-European conquerors had only slightly more children than the local men, R1b lineages would become dominant within a few centuries. Celtic culture lasted for over 1000 years in Continental Europe before the Roman conquest putting an end to the privileges of the chieftains and nobility. This is more than enough time for R1b lineages to reach 50 to 80% of the population.
The present-day R1b frequency forms a gradient from the Atlantic fringe of Europe (highest percentage) to Central and Eastern Europe (lowest), the rises again in the Anatolian homeland. This is almost certainly because agriculture was better established in Eastern, then Central Europe, with higher densities of population, leaving R1b invaders more outnumbered than in the West. Besides, other Indo-Europeans of the Corded Ware culture (R1a) had already advanced from modern Russia and Ukraine as far west as Germany and Scandinavia. It would be difficult for R1b people to rival with their R1a cousins who shared similar technology and culture. The Pre-Celto-Germanic R1b would therefore have been forced to settled further west, first around the Alps, then overtaking the then sparsely populated Western Europe.
The Balkan and Asian branch (Z2103)
Asian Branch
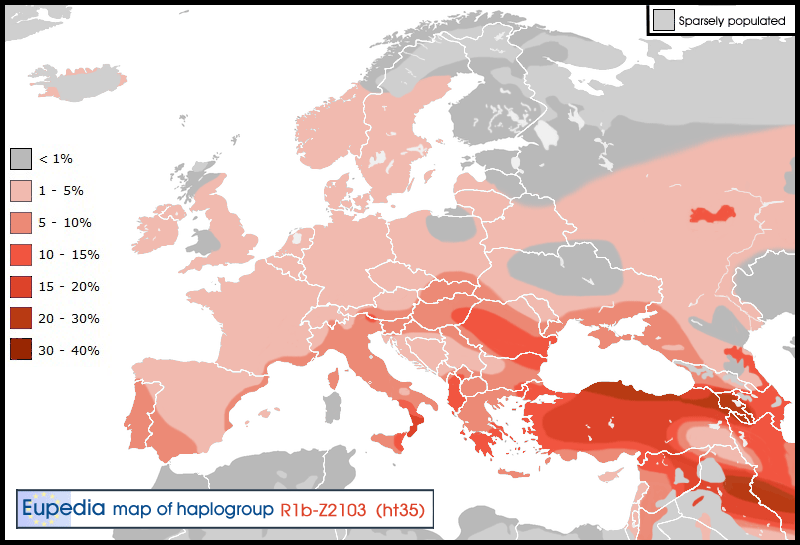
Haak et al. (2015) tested six Y-DNA samples from the eastern reaches of Yamna culture, in the Volga-Ural region, and all of them turned out to belong to haplogroup R1b. Four of them were positive for the Z2103 mutation. In all likelihood, R1b-Z2103 was a major lineage of the Poltavka culture, which succeeded to the Yamna culture between the Volga River and the Ural mountains. It eventually merged with the Abashevo culture (presumably belonging chiefly to R1a-Z93) to form the Sintashta culture. Through a founder effect or through political domination, R1a-Z93 lineages would have outnumbered R1b-Z2103 after the expansion to Central and South Asia, although important pockets of Z2103 survived, notably in Bashkorostan, Turkmenistan and Uyghurstan (Chinese Turkestan).
R1b-Z2103 would have become an Indo-Iranian lineage like R1a-Z93. This is true of two Z2103 subclades in particular: L277.1 and L584. The former is found in Russia to Central Asia then to India and the Middle East, just like the R1a-L657 subclade of Z93. It can be associated with the Andronovo culture and Bactria–Margiana Archaeological Complex, as well as the Indo-Aryan migrations. R1b-L584 is found especially in Iran, northern Iraq, the South Caucasus and Turkey, and correlates more with the Iranian branch of Indo-Europeans, which includes Persians, Kurds and Scythians.
Distribution of Haplogroup R1b-ht35 (Z2103) in Europe
Anatolian Branch
The Hittites (c. 2000-1178 BCE) were the first Indo-Europeans to defy (and defeat) the mighty Mesopotamian and Egyptian empires. There are two hypotheses regarding the origins of the Hittites. The first is that they came from the eastern Balkans and invaded Anatolia by crossing the Bosporus. That would mean that they belonged either to the L23* or the Z2103 subclade. The other plausible scenario is that they were an offshoot of the late Maykop culture, and that they crossed the Caucasus to conquer the Hattian kingdom (perhaps after being displaced from the North Caucasus by the R1a people of the Catacomb culture). In that case the Hittites might have belonged to the R1b-M269* or the R1b-M73 subclade. The first hypothesis has the advantage of having a single nucleus, the Balkans, as the post-Yamna expansion of all Indo-European R1b. The Maykop hypothesis, on the other hand, would explain why the Anatolian branch of IE languages (Hittite, Luwian, Lydian, Palaic) is so archaic compared to other Indo-European languages, which would have originated in Yamna rather than Maykop.
There is substantial archaeological and linguistic evidence that Troy was an Indo-European city associated with the steppe culture and haplogroup R1b. The Trojans were Luwian speakers related to the Hittites (hence Indo-European), with attested cultural ties to the culture of the Pontic-Caspian steppe. The first city of Troy dates back to 3000 BCE, right in the middle of the Maykop period. Troy might have been founded by Maykop people as a colony securing the trade routes between the Black Sea and the Aegean. The founding of Troy happens to coincide exactly with the time the first galleys were made. Considering the early foundation of Troy, the most likely of the two Indo-European paternal haplogroups would be R1b-M269 or L23.
The Phrygians and the Proto-Armenians are two other Indo-European tribes stemming from the Balkans. Both appear to have migrated to Anatolia around 1200 BCE, during the ‘great upheavals’ of the Eastern Mediterranean (see below). The Phrygians (or Bryges) founded a kingdom (1200-700 BCE) in west central Anatolia, taking over most of the crumbling Hittite Empire. The Armenians crossed all Anatolia until Lake Van and settled in the Armenian Highlands. Nowadays 30% of Armenian belong to haplogroup R1b, the vast majority to the L23 subclade.
Most of the R1b found in Greece today is of the Balkanic Z2103 variety. There is also a minority of Proto-Celtic S116/P312 and of Italic/Alpine Celtic S28/U152. L23 could have descended from Albania or Macedonia during the Dorian invasion (see below), thought to have happened in the 12th century BCE. Their language appear to have been close enough to Mycenaean Greek to be mutually intelligible and easy for locals to adopt. The Mycenaean’s might have brought some R1b (M269 or L23) to Greece, but their origins can be traced back through archaeology to the Catacomb culture and the Seima-Turbino phenomenon of the northern forest-steppe, which would make them primarily an R1a1a tribe.
Greek and Anatolian S116 and some S28 lineages could be attributed to the La Tène Celtic invasions of the 3rd century BCE. The Romans also certainly brought S28 lineages, and probably also the Venetians later on, notably on the islands. Older clades of R1b, such as P25 and V88, are only a small minority and would have come along E1b1b, G2a and J2 from the Middle East.
The Great Upheavals circa 1200 BCE
1200 BCE was a turning point in European and Near-Eastern history. In Central Europe, the Urnfield culture evolved into the Hallstatt culture, traditionally associated with the classical Celtic civilization, which was to have a crucial influence on the development of ancient Rome. In Italy, the Terramare culture comes to and end with the Italo-Celtic invasions. In the Pontic steppes, the Srubna culture let place to the Cimmerians, a nomadic people speaking an Iranian or Thracian language. The Iron-age Colchian culture (1200-600 BCE) starts in the North Caucasus region. Its further expansion to the south of the Caucasus correspond to the first historical mentions of the Proto-Armenian branch of Indo-European languages (circa 1200 BCE). In the central Levant the Phoenicians start establishing themselves as significant maritime powers and building their commercial empire around the southern Mediterranean.
But the most important event of the period was incontestably the destruction of the Near-Eastern civilizations, possibly by the Sea Peoples. The great catastrophe that ravaged the whole Eastern Mediterranean from Greece to Egypt circa 1200 BCE is a subject that remains controversial. The identity of the Sea Peoples has been the object of numerous speculations. What is certain is that all the palace-based societies in the Near-East were abruptly brought to an end by tremendous acts of destruction, pillage and razing of cities. The most common explanation is that the region was invaded by technologically advanced warriors from the north, probably Indo-Europeans descended from the steppes via the Balkans.
The Hittite capital Hattusa was destroyed in 1200 BCE, and by 1160 BCE the empire had collapsed, probably under the pressure of the Phrygians and the Armenians coming from the Balkans. The Mycenaean cities were ravaged and abandoned throughout the 12th century BCE, leading to the eventual collapse of Mycenaean civilization by 1100 BCE. The kingdom of Ugarit in Syria was annihilated and its capital never resettled. Other cities in the Levant, Cyprus and Crete were burned and left abandoned for many generations. The Egyptians had to repel assaults from the Philistines from the East and the Libyans from the West – two tribes of supposed Indo-European origin. The Libyans were accompanied by mercenaries from northern lands (the Ekwesh, Teresh, Lukka, Sherden and Shekelesh), whose origin is uncertain, but has been placed in Anatolia, Greece and/or southern Italy.
The devastation of Greece followed the legendary Trojan War (1194-1187 BCE). It has been postulated that the Dorians, an Indo-European people from the Balkans (probably coming from modern Bulgaria or Macedonia), invaded a weakened Mycenaean Greece after the Trojan War, and finally settled in Greece as one of the three major ethnic groups. The Dorian regions of classical Greece, where Doric dialects were spoken, were essentially the southern and eastern Peloponnese, Crete and Rhodes, which is also the part of Greece with the highest percentage of R1b-L23.
Another hypothesis is that the migration of the Illyrians from north-east Europe to the Balkans displaced previous Indo-European tribes, namely the Dorians to Greece, the Phrygians to north-western Anatolia and the Libu to Libya (after a failed attempt to conquer the Nile Delta in Egypt). The Philistines, perhaps displaced from Anatolia, finally settled in Palestine around 1200 BCE, unable to enter Egypt.
Other migrations of R1b
Other migrations occurred from Europe to the Near East and Central Asia during the Antiquity and Middle Ages. R1b-S28 (U152) was found in Romania, Turkey, northern Bashkortostan (a staggering 71.5% of the local population according to Myres et al.), and at the border of Kazakhstan and Kyrgyzstan. Some of it was surely brought by the La Tène Celts, known to have advanced along the Danube, and created the Galatian kingdom in central Anatolia. The rest could just as well be Roman, given that R1b-S28 is the dominant form of R1b in the Italian peninsula. Some have hypothesized that some “lost” Roman legions went as far as Central Asia or China and never came back, marrying local women and leaving their genetic marker in isolated pockets in Asia. A more prosaic version is that Roman merchants ended up in China via the Silk Road, which existed since the 2nd century BCE.
A small percentage of Western European R1b subclades were also found among Christian communities in Lebanon. They are most likely descendants of the crusaders.
4. The Lactase Persistence Allele and R1b Cattle Pastoralists
Lactose (milk sugar) is an essential component of breast milk consumed by infants. Its digestion is made possible by an enzyme, called lactase, which breaks down lactose in simple sugars that can be absorbed through the intestinal walls and into the bloodstream. In most mammals (humans included), the production of the lactase enzyme is dramatically reduced soon after weaning. As a result, older children and adults become lactose intolerant. That is true of a big part of the world population. Some people possess a genetic mutation that allows the production of lactase through adulthood. This is called lactase persistence (LP). Lactase persistence is particularly common among Northwest Europeans, descended from the ancient Celtic and Germanic people, and in parts of Africa where cattle herding has been practiced for thousands of years. The highest incidence for the lactase persistence alleles, known to geneticists as -13,910*T (rs4988235) and -22018*A (rs182549), are found among Scandinavian, Dutch, British, Irish and Basque people. Sub-Saharan populations with lactase persistence have different mutations, such as -14010*C, -13915*G and -13907*G.
R1b men are thought to be the first people on earth to successfully domesticate cattle and to develop a lifestyle based on cattle husbandry and herding during the Pre-Pottery Neolithic (see Neolithic section). Looking for pasture for their cows, R1b tribes migrated from the Near East to the savannah of North Africa (which has since underwent desertification and become the Sahara) and to the Pontic Steppe in southern Russia and Ukraine. For several millennia no other human population was so depended on cattle for their survival as these R1b tribes.
It is known that most Neolithic herding societies consumed at least some animal milk and even made cheese from it (since cheese contains less lactose and is easier to digest for people who are lactose intolerant). In most of Europe, the Middle East and South Asia, people essentially herded goats and sheep, better suited to mountainous environment of the Mediterranean basin, Anatolia and Iran. Goats and sheep could also be kept easily inside villages by sedentary cereal cultivators, while cows needed vast pastures for grazing, which were particularly scare in the Middle East. Domesticated cattle were sometimes found in small number among other Neolithic populations, but the ones that relied almost entirely on them were the R1b tribes of the Pontic Steppe and North Africa. To this very day, semi-nomadic pastoralists in the Sahel, such as the Fulani and the Hausa, who are descended from Neolithic R1b-V88 migrants from the Near East, still maintain primarily herds of cattle. It is among these cattle herders that selective pressure for lactase persistence would have been the strongest.
There has been speculation among geneticists and evolutionary biologists regarding the origin of the lactase persistence allele in Europeans. Over 100 ancient DNA samples have been tested from Mesolithic, Neolithic and Bronze Age Europe and Syria, and the -13910*T allele has been found only in Late Neolithic/Chalcolithic and Bronze Age individuals. The origin of the mutation does not really matter, since it could have been present at low frequencies in the human gene pool for tens of thousands of years before it underwent positive selective pressure among cattle-herding societies. What is certain is that individuals from Bronze Age cultures associated with the arrival of Indo-European speakers from the Pontic Steppe already possessed relatively high percentages of the LP allele. For example the LP allele was found at a frequency of 27% (see Schilz 2006) among the 13 individuals from the Lichtenstein Cave in Germany, who belonged to the Urnfield culture, and were a mix of Y-haplogroups R1b, R1a and I2a2b.
Nowadays, the LP allele is roughly proportional to the percentage of R1b, and to a lower extent R1a, found in a population. In the British Isles, the Low Countries and south-west Scandinavia, where LP is the highest in the world, the combined percentage of R1a and R1b exceeds 70% of the population. In Iberia, the highest percentage of LP is observed among the Basques, who have the highest percentage of R1b. In Italy, LP is most common in the north, like R1b. The lowest incidence of LP in Europe are found in South Italy, Greece and the Balkans, the regions that have the least R1b lineages.
5. R1 Populations Spread Genes for Light Skin, Blond and Red Hair
There is now strong evidence that both R1a and R1b people contributed to the diffusion of the A111T mutation of the SLC24A5, which explains approximately 35% of skin tone difference between Europeans and Africans, and most variations within South Asia. The distribution pattern of the A111T allele (rs1426654) of matches almost perfectly the spread of Indo-European R1a and R1b lineages around Europe, the Middle East, Central Asia and South Asia. The mutation was probably passed on in the Early Neolithic to other Near Eastern populations, which explains why Neolithic farmers in Europe already carried the A111T allele (e.g. Keller 2012, Lazaridis 2014), although at lower frequency than modern Europeans and southern Central Asians.
The light skin allele is also found at a range of 15 to 30% in in various ethnic groups in northern sub-Saharan Africa, mostly in the Sahel and savannah zones inhabited by tribes of R1b-V88 cattle herders like the Fulani and the Hausa. This would presuppose that the A111T allele was already present among all R1b people before the Pre-Pottery Neolithic split between V88 and P297. R1a populations have an equally high incidence of this allele as R1b populations. On the other hand, the A111T mutation was absent from the 24,000-year-old R* sample from Siberia, and is absent from most modern R2 populations in Southeast India and Southeast Asia. Consequently, it can be safely assumed that the mutation arose among the R1* lineage during the late Upper Paleolithic, probably some time between 20,000 and 13,000 years ago.
Fair hair was another physical trait associated with the Indo-Europeans. In contrast, the genes for blue eyes were already present among Mesolithic Europeans belonging to Y-haplogroup I. The genes for blond hair are more strongly correlated with the distribution of haplogroup R1a, but those for red hair have not been found in Europe before the Bronze Age, and appear to have been spread primarily by R1b people.
6. The Maternal Lineages (mtDNA) Corresponding to Haplogroup R1b
Haplogroup R1b is very widespread in most of Europe and across vast swathes of North Africa, the Middle East and Central Asia today. As R1b Indo-Europeans advanced from the Pontic-Caspian Steppe by marrying local women as well as Indo-European women, it is difficult to estimate what were the original mtDNA haplogroups of R1b people back in the steppes, or prior to that in the Neolithic Near East or Paleolithic Eurasia. There are two methods to determine the haplogroups of the original Indo-Europeans.
The first one is to look at ancient DNA. Hundreds of mtDNA samples from Neolithic, Mesolithic and even Paleolithic Europe have been tested to date. Some haplogroups have never been identified outside Northeast Europe before the Bronze Age. These include H2a1, H4, H6, H8, H15, I, T1, U2 and W. All of them except H8 and H15 were found in Bronze Age cultures associated with the diffusion of the Indo-Europeans speakers like Corded Ware and Unetice. Interestingly, Mesolithic and Paleolithic Northeast Europeans were only found to belong to haplogroups U2, U4 and U5, as well as some undetermined subclades of H and T. U4 and U5 were also found in the rest of Europe before the Neolithic. So it could be that mtDNA I, T1 and W came with R1b from the Near East, but this hasn’t been confirmed yet.
It is unclear whether there was any haplogroup V before the Bronze Age either. None could be identified with certainty, but that may simply be because the defining mutations of haplogroup V lie within the coding region of mitochondrial DNA, which is more difficult to recover from ancient samples. Actually not a single V sample has shown up even in Bronze and Iron Age samples. MtDNA V being spread very evenly across all Europe nowadays, it is hard to believe that it wasn’t found among Mesolithic Europeans, and probably Indo-Europeans too.
The problem of the first method is that it only works to identify haplogroups that were brought to Europe by the Indo-Europeans during the Bronze Age, but it doesn’t take into account the haplogroups that the Indo-Europeans had in common with Mesolithic or Neolithic Europeans. For example, it is certain that the Bronze Age Indo-Europeans shared haplogroups U4 and U5 with Mesolithic hunter-gathers, and it is very likely that many H subclades were also present among Mesolithic Europeans, including H1, H3, H10 and H11. The second method attempts to remedy to this issue by looking at the European mtDNA found in Bronze Age and modern samples in parts of Asia that were settled by the Indo-Europeans.
Isolated Indo-European settlements in Asia, such as the Tarim basin in north-west China, the Altai region in southern Siberia, or Bactria and Margiana in southern Central Asia, provide unique opportunities to find European mtDNA in regions that are otherwise genetically Asian. Since there wasn’t any other major European settlement of these regions historically, if we exclude white Russians from the sampled populations, the European mtDNA found in these regions would necessarily correlate with Bronze and Iron Age Indo-Europeans, i.e. haplogroups R1a and R1b. The only interference could come from Middle Eastern mtDNA, especially in Muslim parts of Central Asia and in Xinjiang. Fortunately we have an idea of what mt-haplogroups could have been brought by E1b1b, J1, J2 and T1 people, such as haplogroups HV, N1, J, K, T2 and U3. The Scythians in particular are thought to have hybridized extensively with the descendants of Neolithic southern Central Asians, as ancient Scythian mtDNA contain a lot of these haplogroups (alongside H2a1, U2, U5 and East Asian lineages). But all Scythian Y-DNA so far turned out to be R1a, although they surely included R1b and J2 lineages too.
The European or Middle Eastern lineages found in Central and North Asia include HV, H2a1, H5a, H6, H7, I1, I2, I4, J1b1, J1c (incl. J1c2m), K, N1a, R, T1, T2, U1, U2e, U3, U4, U5, U8, V, W, and X2. If we exclude the haplogroups that are found mostly in R1a countries, or more typical of the Middle East than Europe, what is left as the potential maternal lineages of Bronze Age R1b men are H2a1, H5a, H6, H7, H8, I, J1, K, T1, T2, U4, U5 and V. U3, W and X2 might also be included, although they correlate less with the modern distribution of R1b. The K subclades of likely Indo-European origin include K1a3, K1a4, K1c1 and K2b.
The Indo-European tribes that carried R1b Y-chromosomes were accompanied by other populations assimilated in the Pontic Steppe, or before that in the Caucasus and eastern Anatolia. These minorities included Y-haplogroups G2a3b1, J2b2, R1a-L664 and T1a-L131 and T1a-P77. It is likely that H4, H6, U2 and U4 came from the indigenous steppe population (paternally R1a). Haplogroups H2a1, I1, I2, I4, K1a4 and K1c1, R (mostly R1a), T2 and W are all common in the North Caucasus and could have been assimilated to the R1b population during the Maykop period. Mt-haplogroups H5a, H7, J1c, K, T2 and X2 were all present during the Neolithic in Europe and the Near East. They could have entered the R1b gene pool in the eastern Anatolia or northern Mesopotamia during the Neolithic. R1b tribes are thought to have domesticated cattle in that region 10,500 years ago, yet only moved across the Caucasus some time between 7,500 and 6,500 years ago. For three or four millennia, semi-nomadic R1b herders were bound to have intermingled with some of the Near Eastern or Caucasian neighbours. Another possibility is that these lineages came from Balkano-Carpathian farmers or herders belonging to Y-haplogroups G2a, J2b and T1a who moved to the Pontic Steppe during the Late Neolithic or Chalcolithic and who were absorbed by R1b tribes in the steppes, before the Indo-European migrations took place.
This leaves only mt-haplogroups H8c, H15, J1b1a, U5 and V. as the most likely potential original maternal lineages of R1b tribes before they started mixing with other Near Eastern populations. Looking at deeper subclades, J1b1a and T1a1a display extremely strong correlations with the distribution of Y-haplogroup R1b.
One way of determining what mt-haplogroups R1b tribes carried at the very beginning of the Neolithic, 10,000 years ago, is to compare the above haplogroups with those of African ethnic groups known to possess elevated percentages of R1b-V88. The best studied group are the Fulani, whose mtDNA include three European-looking haplogroups J1b1a, U5 and V making up about 15% of their total maternal lineages. These haplogroups have been identified in all four Central African countries sampled, confirming a strong correlation with haplogroup R1b. Since African R1b-V88 and Eurasian haplogroup R1b-P297 split roughly 10,000 years ago, there is little doubt that J1b, U5 and V were three of the original maternal lineages of R1b people. Only the J1b1b subclade seems to be related to the propagation of Y-haplogroup R1b. Other J1b subclades are geographically restricted to the Near East, particularly from the Caucasus to the Arabian peninsula. J1b might have been the first indigenous Near Eastern lineage assimilated by R1b tribes when they moved into the region (presumably from Russia or Iran, or the Caspian Sea, which was only formed by the melting of Russian glaciers just before the Neolithic began). If that is the case, the Paleolithic R1b people would have belonged exclusively to mtDNA U5 and V, just like the modern Sami, and like a lot of Mesolithic Europeans.